
16.5: Hydrogenation of alkenes and dehydrogenation of alkanes
- Page ID
- 1013

We turn next to reactions in which a hydrogen molecule is added to the double bond of an alkene - forming an alkane - and the reverse, in which H2 is eliminated from an alkane to form an alkene. These reactions, which play an integral role in the metabolism of fatty acids, generally take place on the two carbons adjacent to a carbonyl group.
16.5A: Alkene hydrogenation in fatty acid biosynthesis
In the final step of the fatty acid synthesis cycle, a double bond between Cα and Cβ is reduced to a single bond by hydrogenation.
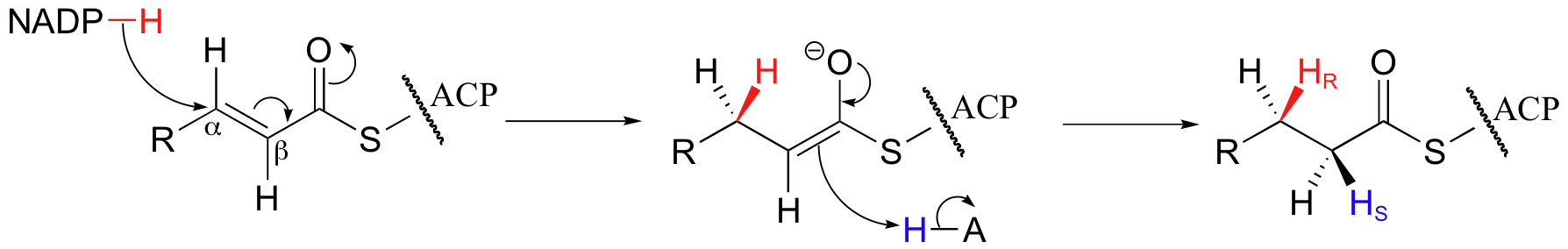
Mechanistically, this can be thought of as a Michael addition with a hydride nucleophile. NADPH acts as the hydride donor. Stereochemically, it is a syn addition (the protons labeled HR and HS in the product are the ones that have been added).
Next, let us consider the complimentary dehydrogenation reaction in fatty acid degradation, in which a double bond is introduced between the Cα and Cβ carbons, with concurrent loss of a hydride and a proton (it is HR on both Cα and Cβ that are lost).
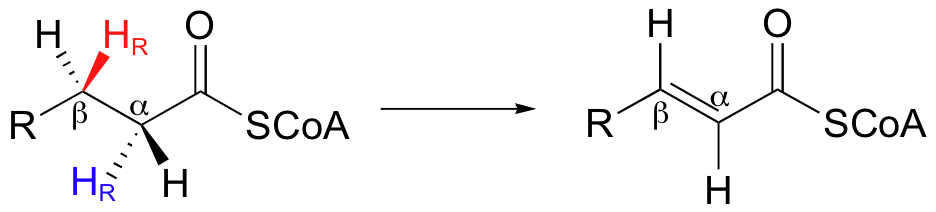
Notice that this reaction is not the exact reverse of the hydrogenation reaction in fatty acid biosynthesis. First, the thioester linkage is to coenzyme A as opposed to an acyl carrier protein (ACP). Secondly, the stereochemical details are clearly different (the hydrogenation is syn, the dehydrogenation is anti). Third, and most importantly for this discussion, while the hydride donor in the hydrogenation reaction is NADPH, the hydride acceptor in the dehydrogenation reaction is not NAD+ or NADP+ - rather, it is a flavin coenzyme.
16.5B: The flavin coenzymes
There are two forms of flavin coenzyme used by 'flavoenzymes': flavin adenine dinucleotide (FAD) and flavin mononucleotide (FMN), which is lacking the AMP unit found in FAD.
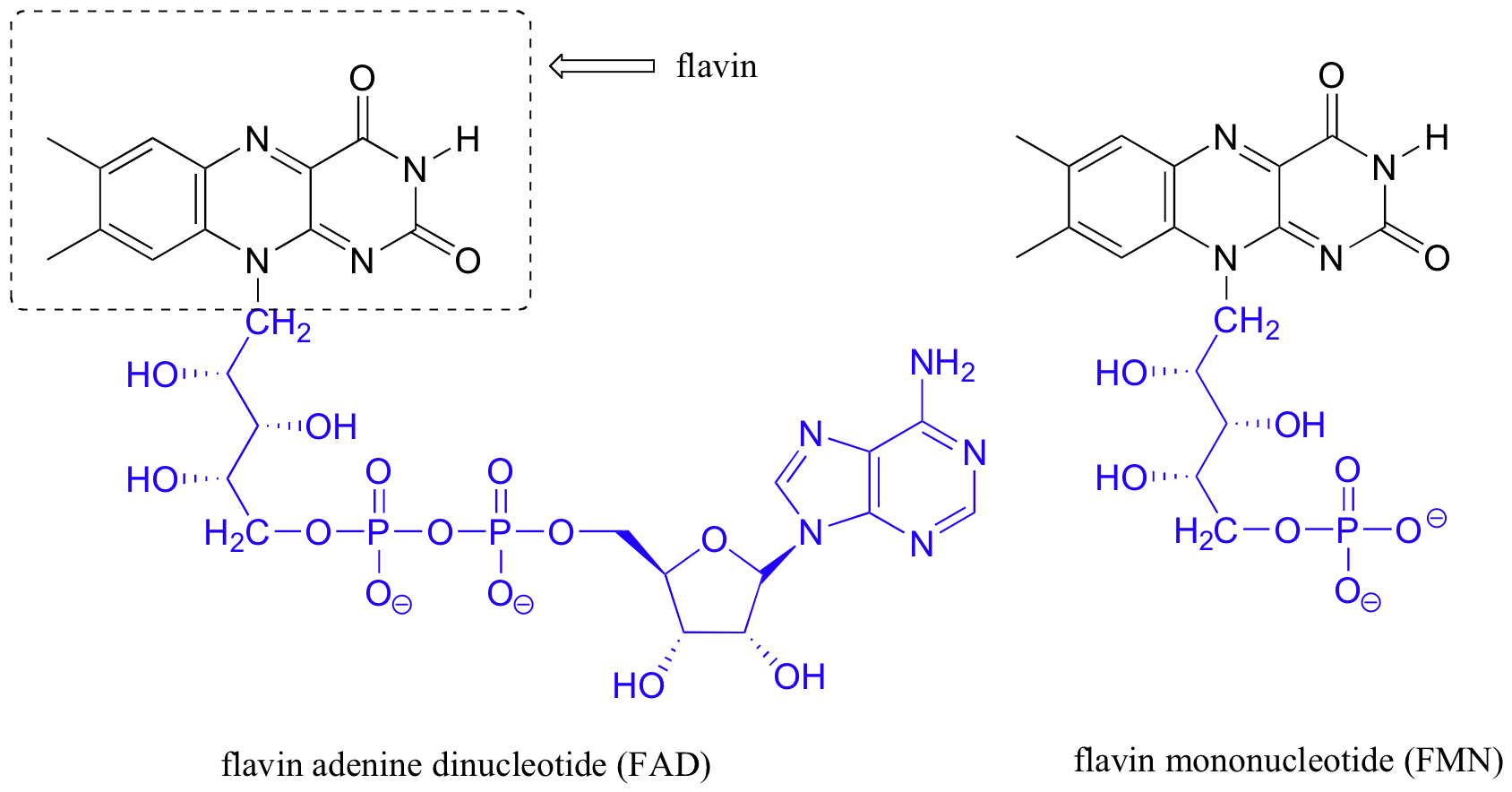
Because the redox-active part of both coenzymes is the flavin group, both FAD and FMN are generally depicted with an R group designating the remainder of the molecule.
The reduced (hydrogenated) forms of these cofactors are abbreviated FADH2 and FMNH2.
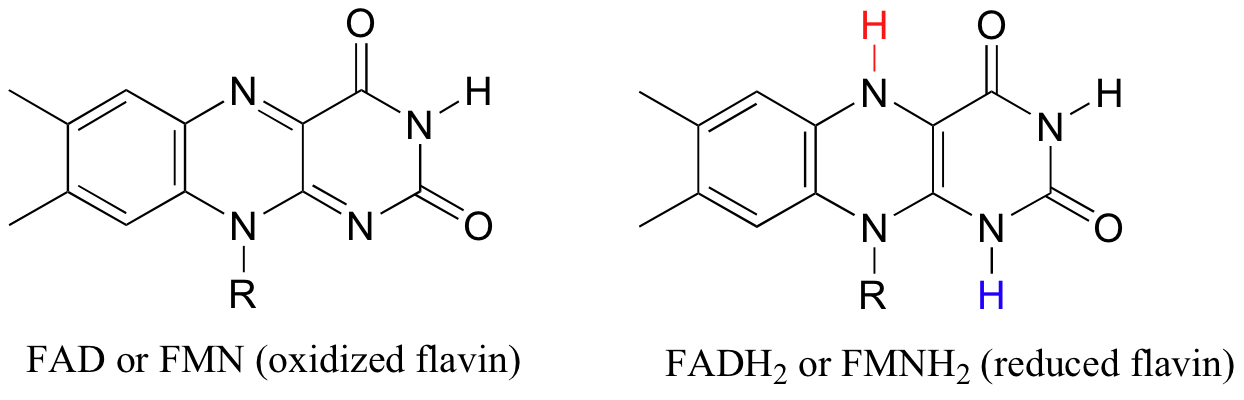
The flavin coenzymes are synthesized in humans from riboflavin (vitamin B2), which we obtain from our diet (the structure of riboflavin is the same as that of FMN, except that riboflavin lacks the phosphate group). The flavin system absorbs light in the visible wavelengths and has a distinctive deep yellow color - it is riboflavin, and to some extent FAD and FMN, that give urine its color.
Like NAD(P)+, FAD and FMN are able to accept a hydride ion (and a proton), and FADH2 and FMNH2 in turn can serve as hydride donors in hydrogenation reactions.

Unlike the nicotinamide cofactors, however, flavins are also able to function in single electron transfer (radical) mechanisms, as well as in many "oxygenase" reactions in which organic molecules are oxidized by the addition of one or more oxygen atoms. The role of FAD and FMN in these types of mechanisms will be discussed in sections 16.10A and 17.3C. For now, we will concentrate on the role of flavin in 'two electron' redox reactions involving direct hydride transfer.
16.5C: Alkane dehydrogenation in fatty acid degradation
In the dehydrogenation step of fatty acid degradation, we can see that a trans double bond forms between Cα and Cβ when the fatty acid substrate loses a hydride and a proton.
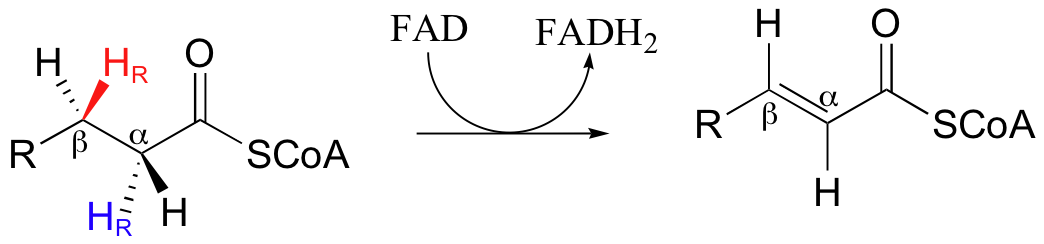
It has been established experimentally that the first step in this reaction is proton abstraction at Cα.

What happens next is not yet entirely clear - it certainly makes sense to propose a simple E1cb elimination of the hydride on Cβ, with FAD serving as the hydride acceptor.

Because FAD also is capable of single-electron chemistry, however, an alternative radical mechanism (which we will examine in section 17.3C) is also possible. Which mechanism is actually occurring is still a matter for debate.
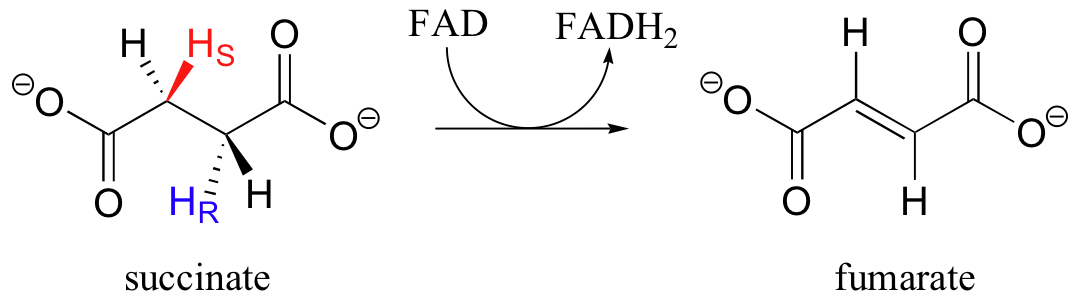
A very similar anti alkane dehydrogenation reaction occurs as part of the citric acid (Krebs) cycle, when succinate is oxidized to fumarate and FAD is reduced to FADH2.
16.5D: More examples of enzymatic alkene hydrogenation
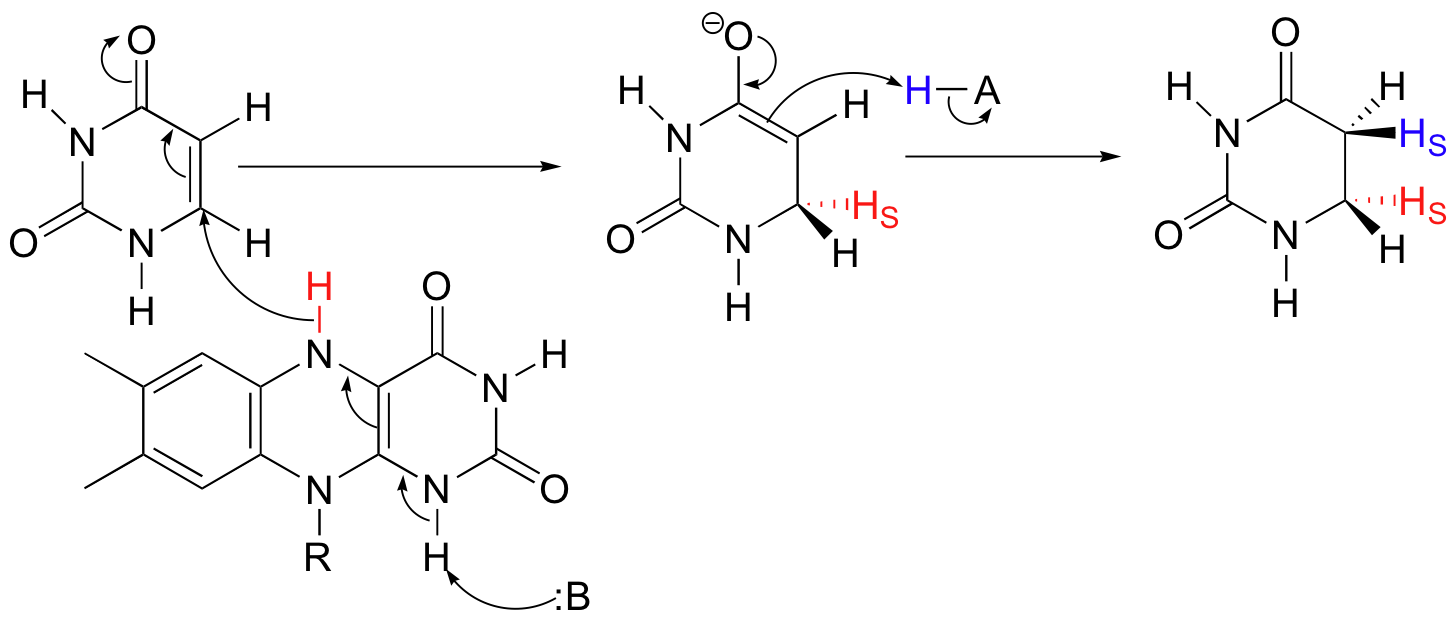
Alkene hydrogenation reactions can involve either reduced flavin coenzyme or NADPH as the direct source of hydride. Breakdown of the RNA nucleotide base uracil begins with the transfer of a hydride from FMNH2 to a conjugated alkene.
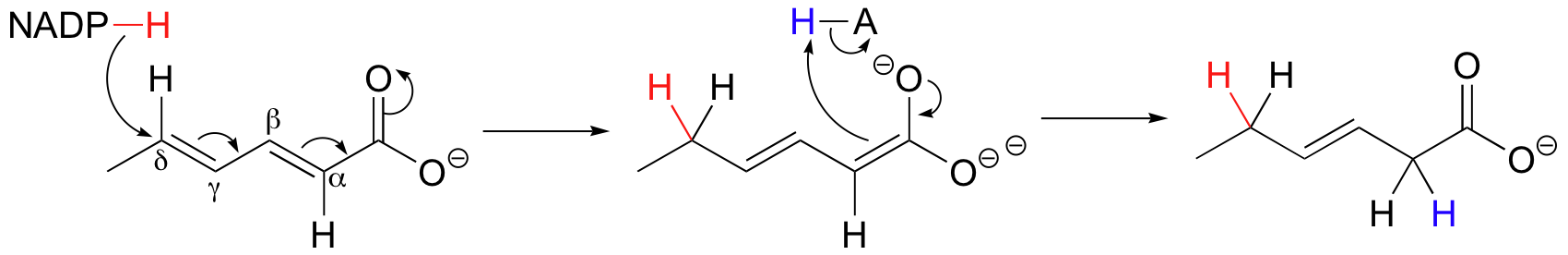
While most alkene hydrogenations involve double bonds in the α-β position relative to a carbonyl, conjugated γ-δ and γ-α hydrogenation is also possible. Below is an example of NADPH-dependant, γ-α addition of hydrogen to a polyunsaturated thioester (J. Biol. Chem 2005, 280, 3068).
Bilirubin, the molecule responsible for the yellowish color of bruises, is formed from the hydrogenation of a double bond in biliverdin, which is a product of heme degradation (heme is an iron-containing coenzyme in the oxygen-carrying blood protein hemoglobin).

There are also some examples of enzymatic hydrogenation of isolated (non-conjugated) double bonds, such as this example in the biosynthesis of chlorophyll (Eur. J. Biochem 1998, 251, 413).
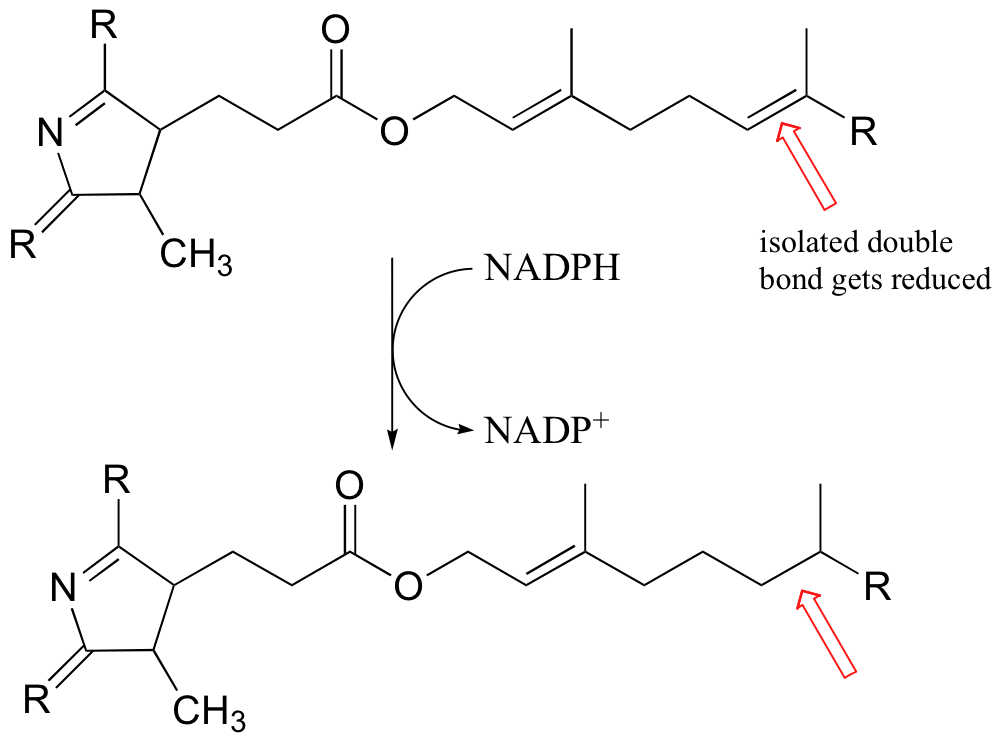
The mechanism for this reaction is not yet known.