
16.2: The importance of redox reactions in metabolism
- Page ID
- 1010

An understanding of the redox chemistry of organic compounds is central to understanding the metabolism of living things. Think back to the electrochemistry that you learned in general chemistry. In your laboratory class, you may have set up a galvanic cell consisting of a copper electrode immersed in an aqueous copper nitrate solution, connected by a wire to a zinc electrode immersed in an aqueous zinc nitrate solution. When you completed the cell with a salt bridge, an electrical current began to flow - what you constructed was a simple battery. Over time, the copper electrode got heavier as metallic copper was deposited on the copper cathode, while the zinc anode slowly dissolved into solution. The redox reaction that was occurring was:
Cu+2(aq) + Zn(s) → Cu(s) + Zn+2(aq)
Electrons were flowing from zinc metal to copper cations, creating zinc cations and copper metal. You could have predicted beforehand that the spontaneous flow of electrons would go in this (and not the opposite) direction just by looking at a table of standard reduction potentials (such a table is was no doubt in your general chemistry text). Zinc metal is a stronger reducing agent than copper metal, meaning that, under identical conditions, more energy is released by oxidizing one mole of zinc metal to Zn2+ than is released by oxidizing a mole of copper metal to Cu2+. Therefore, the transfer of two electrons from zinc metal to Cu2+ is a thermodynamically downhill process, whereas the transfer of two electrons from copper metal to Zn2+ is thermodynamically uphill.
Let's try to extend the idea of redox reactions to a organic/biochemistry context. When we drink a can of cola, we are taking in fuel molecules mainly in the form of the disaccharide sucrose, which is a form of sugar we quickly convert to glucose. Then, in a long series of enzyme-catalyzed reactions, we 'burn' the glucose molecules: we transfer electrons from glucose to molecular oxygen, oxidizing the carbon molecules to carbon dioxide and reducing O2 to water:
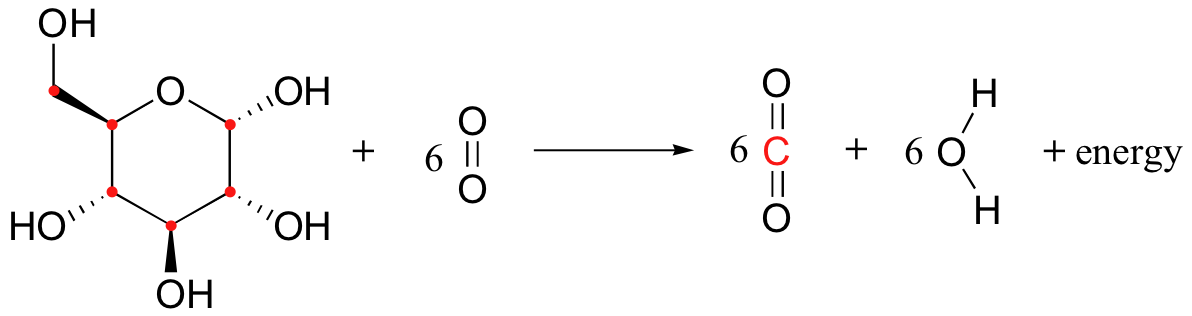
Glucose is a much stronger reducing agent than water (or, to put it in a more conventional way, O2 is a much stronger oxidizing agent than CO2). Therefore, the transfer of electrons from glucose to O2 is a thermodynamically downhill, energy-releasing process, just like the transfer of electrons from zinc metal to copper ion. And while you could have used the energy released by the zinc/copper redox reaction to light a small light bulb (or to give your lab partner a nice little shock!), your cells use the energy released by the glucose/oxygen redox process to carry out a wide variety of energy-requiring activities.
Example 16.3
If the oxidation of glucose to CO2 is thermodynamically downhill, why doesn't a teaspoon of corn sugar (pure glucose) spontaneously oxidize before your eyes?
In your general chemistry copper/zinc battery, was it possible to get the reaction to run in the uphill direction - to oxidize copper and reduce zinc?
Zn+2(aq) + Cu(s) + energy → Zn(s) + Cu+2(aq)
Of course it was! All you had to do was to provide energy in the form of an external electrical current (you do something like this every time you recharge a battery). In the same way, the photosynthesis systems of plants are able to reduce carbon dioxide and oxidize water to form glucose and molecular oxygen.
6CO2 + 6H2O + energy → C6H12O6 + 6O2
Because water is such a weak reducing agent, this is very much an 'uphill' reaction thermodynamically, and therefore it requires the input of energy from an external source - the sun. On a global scale, the breakdown and oxidation of sugar to CO2 by non-photosynthetic organisms (like people) and the subsequent reduction and condensation of CO2 back to sugar and oil by plants is a central part of what ecologists refer to as the 'carbon cycle'.
Like plants, animals can also drive some kinds of unfavorable, 'uphill' redox reactions with energy input from an external source . But where plants use energy from the sun, we use the energy we get from oxidizing food molecules to drive our uphill redox reactions - this is why we need to eat in the first place! Also unlike plants, we are not able to make glucose out of carbon dioxide - we don't have the right enzymes.
In general the more reduced an organic molecule is, the more energy is released when it is oxidized to CO2. Going back to our single-carbon examples, we see that methane, the most reduced species, releases the most energy when oxidized, while formic acid releases the least:

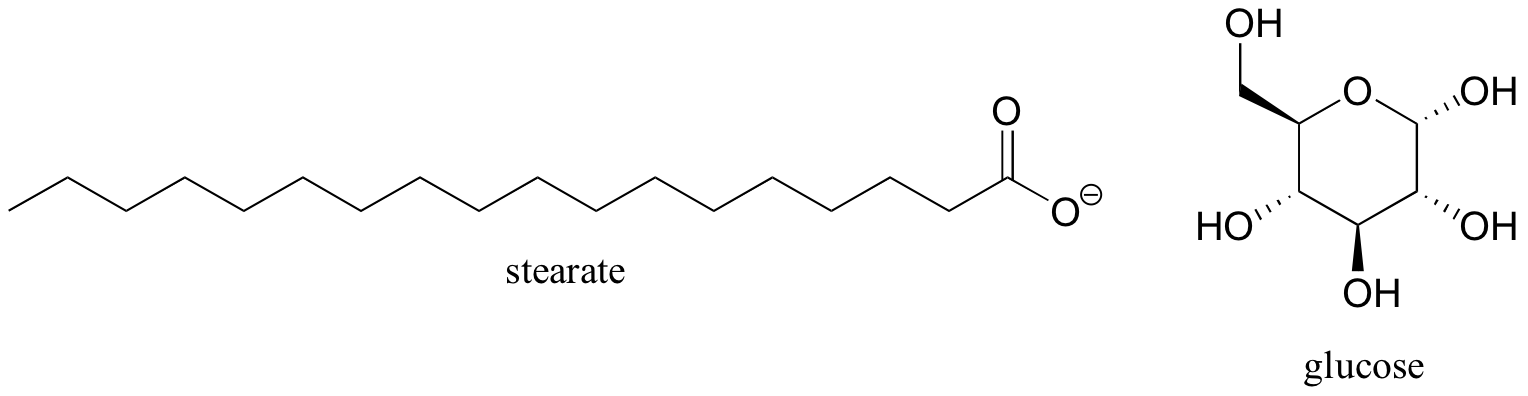
This explains why fatty acids (like stearate, below), in which most of the carbons are in the alkane oxidation state, contain more energy than sugars, where most of the carbons are in the alcohol oxidation state.
Example 16.4
Which carbon in the glucose molecule is not in the alcohol oxidation state? What state is this carbon in?
Example 16.5
In general, which would you expect to contain more calories per gram, saturated or unsaturated fat?
Example 16.6
Which would you expect to contain the most and least calories: a bottle of grape juice, the wine made from the fermentation of the grape juice, or the vinegar that was produced when the wine was left out uncovered for a few weeks? Explain. (Hint - a wine-maker can tell that his grape juice is fermenting because it produces bubbles).
After we break down and oxidize sugar and fat molecules to obtain energy, we use that energy to build large, complex molecules (like cholesterol, or DNA) out of small, simple precursors (like acetyl-CoA or pyruvate - unlike plants, we can't start with CO2). The biosynthetic process requires the input of energy in large part because it is reductive: the carbons in the large products are in a reduced state compared to the small precursors (just look at the structure of cholesterol compared to that acetyl-CoA, from which it is derived - in which molecule are the carbons, on average, more reduced?).
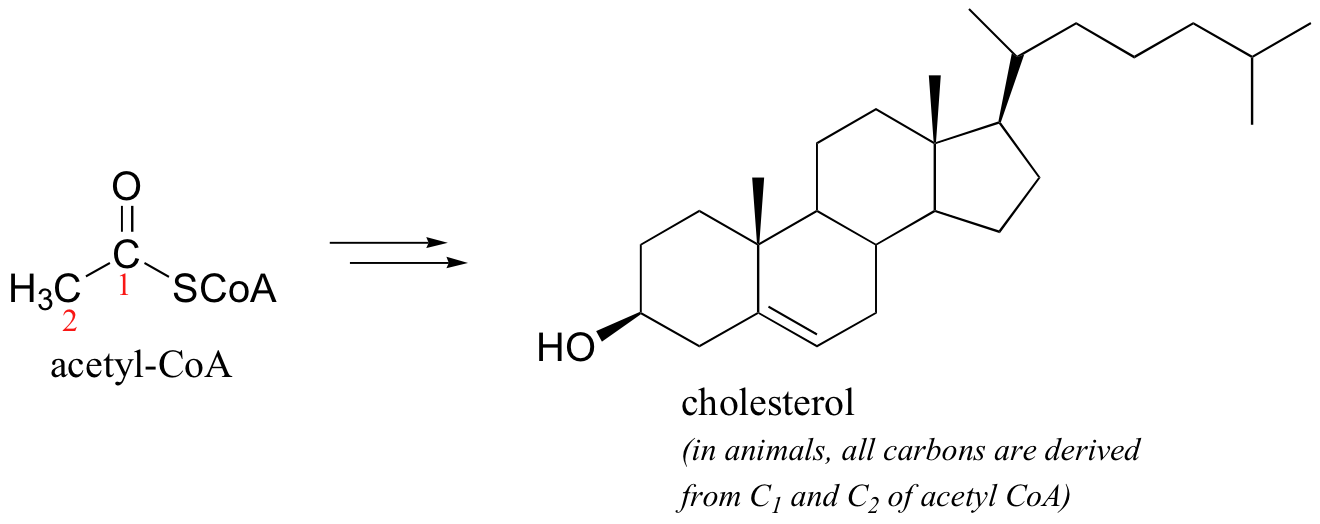
Overall, you can look at animal metabolism as a process in which reduced, high energy organic (food) molecules are oxidized to obtain energy, which is then harnessed in part to drive the uphill, reductive synthesis of our own high energy organic molecules.